
 朗读本文
朗读本文1884年,法国博物学家Künckel d'Herculais描述了来自越南北部下龙湾附近的长臂猿新种,也就是现在的东黑冠长臂猿(Nomascus nasutus),它隶属于灵长目(Primates),长臂猿科(Hylobatidae),冠长臂猿属(Nomascus),是全球最濒危的25种灵长类之一,被世界自然保护联盟(IUCN)红色名录列为全球极度濒危物种,我国一级重点保护野生动物。该物种历史上曾广泛分布于红河以东(中文名中“东”字的由来)的中国南部和越南北部。由于偷猎和栖息地的丧失,该物种的种群数量急剧下降,自20世纪50年代起一度认为已经从中国灭绝,60年代后越南也没有该物种分布的确切消息。幸运的是,调查人员分别于2002年和2006年在越南高平省重庆县和中国广西壮族自治区靖西市的喀斯特森林中重新发现了该物种。“消失”近半个世纪的东黑冠长臂猿重回大众视野也一度成为国际保护生物学界轰动一时的科学发现。
东黑冠长臂猿的重新发现给了人类重新认识和了解它们的机会。自2007年,由中山大学范朋飞教授带领的科研团队在中国靖西与越南高平交接的一小片喀斯特森林中开始对东黑冠长臂猿的觅食、过夜、繁殖、鸣声、社会系统、保护等方面展开长期研究。现在人们对东黑冠长臂猿的了解基本源于他们对3-5个家庭群日复一日的观察和记录。

东黑冠长臂猿(赵超 摄)
臂展可达一米多,雄全黑、雌黄棕色东黑冠长臂猿的腿很短,但手臂极长,其臂展可达一米多,主要以双臂交替摆动和攀援的方式在林间移动。成年个体具有明显的毛色性二型,成年雄性全身黑毛,而成年雌性的毛色更鲜艳,以黄棕色为主,成年雌性头顶的黑色冠斑可从肩部延伸至背部,脸部有一圈非常明显的白毛。基于它们历史上曾广泛分布于红河以东和成年雌性头顶的黑色冠斑,由此得名东黑冠长臂猿。它们的婴猿全身黑毛,雄性个体从出生到成年一直都是身披黑毛,而雌性个体在接近成年时转变为黄棕色。然而,冠长臂猿属其它物种的新生婴猿毛发呈奶白色,在一到两周内变成和母亲颜色接近的黄棕色,在一岁左右无论雌雄都逐渐转变成黑色,而雌性在接近成年时则再次转变成黄棕色。总体而言,雄性东黑冠长臂猿不会经历毛色变化,雌性东黑冠长臂猿经历一次毛色变化。而冠长臂猿属其它物种的雄性经历两次毛色变化,雌性则经历三次毛色变化。这样的差别是东黑冠长臂猿作为一个独立物种的有力证据之一。

东黑冠长臂猿(黄嵩和 摄)

东黑冠长臂猿(黄嵩和 摄)
一生几乎不下地,主要取食植物东黑冠长臂猿是典型的树栖型动物,一生几乎不下地,中越边境上这片约30平方公里的喀斯特森林是它们最后的庇护所。该区域是典型的喀斯特石灰岩景观,由峰丛和峰谷组成,峰丛上裸露的岩石陡峭且锋利。由于烧炭,薪柴砍伐,农业耕种,放牧等历史因素,当前该区域主要由一些次生林组成,乔木的平均高度只有10.5米,这远远低于其它长臂猿栖息地中乔木的高度。该区域是典型的季风性气候,具有明显的季节性变化。1月是该区域均温最低的月份,9月是均温最高的月份;雨季从5月开始,持续到10月,这一时期果实丰富;从11月至次年4月,降雨减少,果实相对匮乏。
研究发现,东黑冠长臂猿取食81种植物和少量动物,尤其偏好取食大叶水榕(Ficus glaberrima)、大青树(Ficus hookeriana)和小叶榕(Ficus microcarpa)的果子。藤本植物也是它们的重要食源植物,5-9月间它们取食毛脉崖爬藤(Tetrastigma pubinerve)的果,全年取食其嫩叶。东黑冠长臂猿具有明显的觅食季节性策略。在雨季,东黑冠长臂猿也会优先取食果实,花超过60%的时间取食果实,仅仅花不到2%的时间取食叶和芽。然而在旱季,取食叶和芽的时间超过50%,其中3月是取食叶和芽的顶峰,甚至可达到95%。总体而言,果实占据东黑冠长臂猿全年食谱的52%,接下来依次是35%的叶和芽,9%的动物性食物。东黑冠长臂猿非常节俭,不浪费食物,取食时它们只摘成熟的果实填饱肚子,而不会破坏那些未成熟的果子。与生活在云南无量山的西黑冠长臂猿相类似,东黑冠长臂猿偶尔也会捕食松鼠这样的小型哺乳动物。

东黑冠长臂猿(马长勇 摄)
早睡早起,傍晚回过夜树过夜东黑冠长臂猿是昼行性动物,习惯早睡早起。每天日出之后,东黑冠长臂猿就会离开过夜树开启一天的活动,日落前一个半小时会进入过夜树。它们偏好选择一些高大的乔木过夜,躲避天敌、避风保温是影响它们选择过夜树的主要因素。一个家庭往往拥有数百棵不同的过夜树,有些过夜树会被反复利用数十次,但是它们很少连续两个晚上利用同一棵过夜树。过夜时,一个家庭中的不同个体往往分树而眠,未成年个体习惯跟随自己的母亲在一起休息。它们每天待在过夜树上的时间长达13-14小时,养成了非常好的卫生习惯,通常会离开过夜树后排便。为了促进彼此之间的感情,维持融洽的家庭关系,群内的个体还会经常在过夜树上梳理毛发。
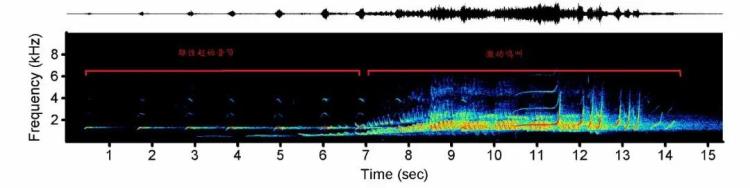
传递信息,雄雌发出美妙二重唱东黑冠长臂猿是喀斯特森林中的歌唱家,家庭群中的成年雄性和成年雌性相互配合能够发出美妙的二重唱。二重唱从成年雄性的起始音节、简单的重复音节开始,随着鸣叫的深入,雄性叫声序列中的简单重复音节数量增加,调节音节的结构变得复杂,在高潮部分,雌性发出激动鸣叫与雄性配合形成激动鸣叫序列。它们的歌声极具穿透力,可以传播1-2公里,也能够传递“我是谁”的信息,经验丰富的研究人员往往可以根据二重唱辨别不同的家庭群。在一个家庭群内,成年个体之间会形成具有一定规律和固定模式的重唱,这些重唱被推断为具有防卫领域、防卫食物资源、吸引配偶、强化配对关系、凝聚群体、调节群间关系等作用。在防卫领域时,东黑冠长臂猿偏好在家域的中心位置鸣唱,这被认为是对喀斯特环境的适应,以便于更好地传递声音中携带的信息。
东黑冠长臂猿的鸣唱具有明显的二级结构,一次鸣叫由很多相似的句子组成,每个句子则包含几种不同的音节,这些音节有相对固定的排列方式。通过多年的研究发现,雄性东黑冠长臂使用越多的音节的持续时间越短,句子越长时其包括的音节则越短,非常符合人类语言的Zipf简约定律(通常使用越频繁的词,其持续时间越短)和Menzerath定律(句子越长时其包括的短语和从句则越短),这或许将有助于我们找到更多人类语言进化的线索。
范鹏来/提供
家庭群一雄二雌,后代8岁离家东黑冠长臂猿生活在稳定的一雄二雌家庭群中,群内两只雌性都能繁殖,群内雌性之间关系融洽,经常会相互理毛,在一起取食,很少发生冲突。成年雌性一般3年生一胎,当前还没记录到双胞胎的现象。两只雌性的未成年后代也会生活在群内,群体大小一般在5-9只之间。婴猿在4个半月龄时才会离开母亲开始独自活动,8月龄时开始取食固体食物,11月龄时开始给其它家庭成员理毛,24月龄时断奶。8岁左右,两性后代都可能会从原生家庭迁移,但雄性离家的可能性高于雌性。
“杀婴行为”普遍存在于灵长类动物之中,是指新上位的成年雄性有时候会杀死群中存在的婴幼个体,促使雌性快速进入发情期,以便新任雄性能够在最短的时间内获得自己的后代。长期的野外观察发现,每当东黑冠长臂猿家庭群的成年雄性更替时,前任雄性的婴猿后代往往凶多吉少。被取代后的前任雄性会离开原来的群体,不知所踪。
适宜栖息地受到严格保护自东黑冠长臂猿被重新发现以来,我国政府高度重视其保护状况,在野生动物盗猎、森林防火、项目管理、公众保护教育等领域开展了很多工作。范朋飞教授带领的团队在一线长期的研究极大地促进了政府、社会公众、科学界对该物种的认识。2009年,我国政府在广西靖西市建立了广西邦亮长臂猿自治区级自然保护区,2013年升级为广西邦亮长臂猿国家级自然保护区。当前,东黑冠长臂猿所有的适宜栖息地都受到法律严格保护,保护区内禁止烧制木炭、薪柴砍伐、盗猎。保护区管理局也组建了一支队伍定期开展种群监测和日常巡护,自2006年在我国重新发现该物种以来,已经完全杜绝了其被偷盗猎。2016年中越两国政府、科学家联合开展了同步调查,共记录到22群136只东黑冠长臂猿。其中,5群33只生活在我国广西邦亮长臂猿国家级自然保护区内。虽然其种群数量呈现出缓慢增长的模式,但东黑冠长臂猿依然是仅次于海南长臂猿的世界第二濒危的类人猿物种。

东黑冠长臂猿(赵超 摄)
种群复壮仍需突破困境虽然东黑冠长臂猿得到了社会各界广泛关注,但它们的保护之路依然任重道远。目前,东黑冠长臂猿生活在孤岛状的一小片喀斯特森林中,周围被河流、农田和村庄隔离。结合遥感和地理信息系统技术的分析发现,该区域适宜东黑冠长臂猿栖息的高质量生境区域最多只能够承载20群长臂猿生活。即使算上一些低质量的栖息,最多也只能承载26群长臂猿生活。令人担忧的是,当前的种群数量已经非常接近环境容纳量,因此亟需采取相应的措施扩大它们的栖息地。由于长臂猿的重要食物树在喀斯特环境中自然恢复的周期长,所以有必要人为种植一些重要食物树以加速森林恢复。其次,两块潜在的栖息地都不在保护区范围内,应加大这些区域的保护力度,严格禁止在这些区域放牧、种植和砍伐薪柴。第三,东黑冠长臂猿仅仅只分布在中越边境这片狭长的喀斯特森林中,抵御自然和人为灾害的能力依然非常弱。在全球新冠疫情形势依然严峻的态势下,建议相关管理部门采取最严格的保护管理措施,禁止非必要人员进入保护区,最大限度地降低东黑冠长臂猿的死亡风险。最后,有关东黑冠长臂猿的很多科学问题依然有待探索和揭秘,深入开展科学研究依然是制定合理的保护方案和政策的最佳指导依据,从而促进东黑冠长臂猿的种群复壮。(信息来源 中国野生动物保护协会 编辑 王强)





























 渝公网安备50010702502703号
渝公网安备50010702502703号