
 朗读本文
朗读本文
棘龙 又称为棘背龙、脊背龙,是种兽脚亚目恐龙,生存于白垩纪的非洲北部,约为1亿1400万年到6500万年前。最明显的特征是背部有明显的长棘,是由脊椎骨的神经棘延长而成,长度可达2.2米,长棘之间推断有皮肤连结,形成一个巨大的帆状物。对于这帆状物的功能,科学家们众说纷纭。
模式种

埃及棘龙
Spinosaurus aegyptiacus
棘龙属目前有两个已命名种,分别为:
埃及棘龙 S. aegyptiacus
摩洛哥棘龙 S. marocannus

摩洛哥棘龙的骨架模型
棘龙的第一个化石是在1912年发现于埃及西部的拜哈里耶绿洲,并由德国古生物学家恩斯特·斯特莫在1915年命名为模式种埃及棘龙。最初的标本在第二次世界大战期间被摧毁,但最近几年发现了其他的头颅骨、爪子、胳膊、尾锥、头冠等身体部分的化石。摩洛哥棘龙是在1996年由戴尔·罗素所研究、命名,罗素是根据颈椎长度而将它们建立为一个新种。罗素比较颈椎椎体的长度、椎体后缘的高度,发现埃及棘龙的比例为1.1,而摩洛哥棘龙的比例为1.5。然而,数名研究人员认为摩洛哥棘龙的颈椎长度只是个体间的变化,而埃及棘龙的正模标本已被轰炸破坏,因此无法比对两者是否为相同动物,而且无法确定摩洛哥棘龙的颈椎是第几节。
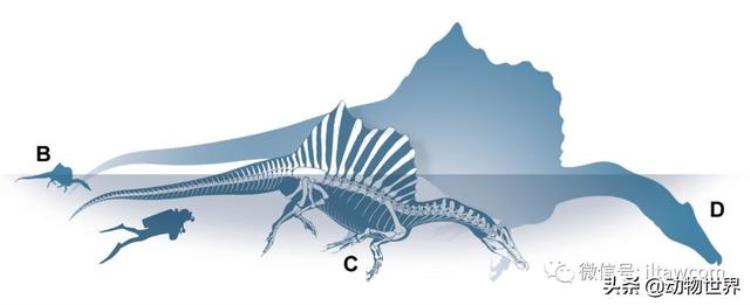
三个不同棘龙个体与人类的体型比较,从左到右分别为幼年个体(MSNM V6894,迄今发现最小的个体)、新模标本(FSAC-KK18888)与迄今发现最大的个体(MSNM V4047)
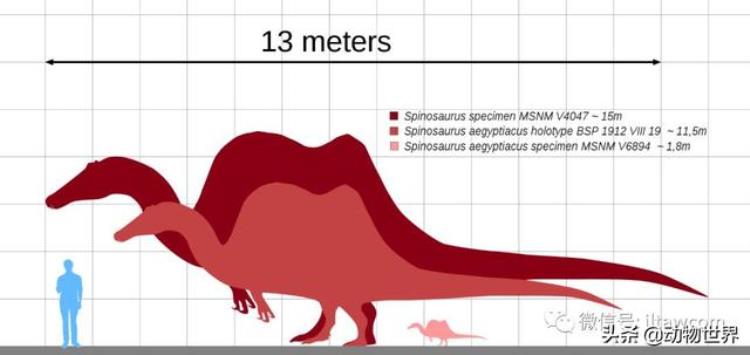
根据最近的估计,棘龙是目前已知最大型的陆生肉食性恐龙之一,长度远远大于暴龙等传统印象中的凶猛恐龙,但是重量却和暴龙持平或者低于它,身高普遍低于大型兽脚类掠食者。
这些推算显示棘龙身长13到18米,高度约为2.1至4.8米,体重为4到18吨。

自从棘龙被发现之后,它们成为最长、最大型兽脚类恐龙的候选者之一,但以前一般大众对于棘龙的巨大体型并没有概念,直到棘龙出现在2001年的电影《侏罗纪公园 III》,以及2005年对于它们新标本的研究。弗雷德里克·冯·休尼博士与数十年后的唐诺·葛勒特都在他们的研究中,将棘龙列为最巨大的兽脚类恐龙之一。他们认为棘龙身长为15米,重量为6吨以上。

在1988年,格里高利·保罗也将棘龙列为最长的兽脚类恐龙,身长一样被叙述为15米,但估计的体重数据较低,约4吨。


达鲁·沙索发现的埃及棘龙的部分上颌,编号MSNM V4047
该标本由前上颌骨、部分上颌骨、以及部分鼻骨所构成,这些部位总长度为98.8公分。头颅骨被估计有2.15米长。该标本被估计约18到20米长,重量约23吨。
在2005年,克里斯提亚诺·达鲁·沙索认为棘龙、似鳄龙有类似的身体与头颅骨比例,而似鳄龙的身长估计约为11米,体重约3.8吨。达鲁·沙索等人根据这个数据,而将棘龙身长估计为18到20米,体重为20到23吨。之后有极少数研究人员也质疑,棘龙科的不同属是否有不同的头颅骨形状,而这将导致不同的身长估计值。

棘龙的口鼻部较窄,布满笔直的圆椎状牙齿,牙齿缺乏锯齿边缘,类似其他的棘龙科恐龙。每块前上颌骨有6或7颗牙齿,每块上颌骨有12颗牙齿。前上颌骨的第2、第3颗牙齿明显地较大,长度13厘米,而和骨后方的一些牙齿略小,但也有7厘米长。


前上颌骨牙齿、上颌骨的大型牙齿间形成一个缺口。此缺口与下颌的大型牙齿吻合。棘龙的口鼻部前端略为膨大,眼睛前方有一个小型突起物。
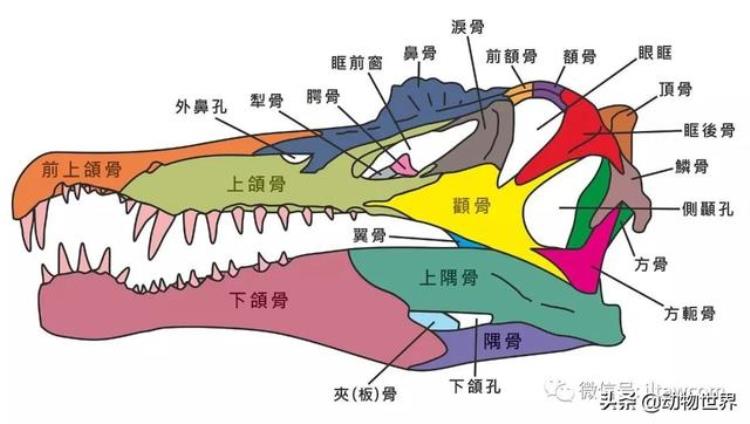

在2005年,克里斯提亚诺·达鲁·沙索根据三个标本,并参考激龙的头颅骨后半段形状,而估计棘龙的头颅骨长度约为2.15米。之后有少数研究人员质疑,棘龙科的不同属是否有不同的头颅骨形状。

棘龙的背部有明显的长棘,是由脊椎骨的神经棘延长而成,长度可达2.2米,长棘之间推断有皮肤连结,形成一个巨大的帆状物。然而有些科学家认为这些长棘是由肌肉覆盖着,形成隆肉或是背脊。
棘龙拥有独特的帆状物,早在1915年,恩斯特·斯特莫就认为这些背部结构物支撑著脂肪构成的隆肉。在1997年,Jack Bowman Bailey最近提出棘龙、豪勇龙的背棘并非细棒,而是前后轴宽广,不同于盘龙目的扁平背帆,而是类似巨角犀、长角野牛的背脊,所以棘龙的背棘应是支撑着较厚、较肥胖的结构物,而非细的帆状物。

本人(仅个人观点)赞同以上的观点,背棘起着支撑作用。那么它们的形体将有可能发生大的变化,更倾向于躯体的粗壮,也因此在生态习性的推测方面也会发生巨大的改变。

它们的体型也许会变成这样

棘龙的帆状物功能仍未确定:科学家们已提出数个假设,包含调节体温、吸引异性的展示物。此外,如此明显的背部特征可使棘龙的外表看起来比较大,可威胁其他动物。

如果这些帆状物拥有大量的血管,棘龙可使用帆状物的大型表面积来吸收热量。这显示棘龙至少是某种程度的温血动物,并生存于夜间气温低、冷,但天空通常很少云的地区。棘龙与豪勇龙被认为为生存于早期撒哈拉沙漠的边缘,这可解释上述的功能。
这些帆状物也有可能用来释放多余的体温,而非收集热量。根据霍尔丹法则,与身体体积相比,大型动物反而拥有较小比例的表面积,它们释放的热量温度较高,而吸收的热量温度较低。这些恐龙的帆状物增加相当程度的身体皮肤面积,而且所增加体积最少。此外,如果将这些帆状物远离太阳方向,或是以90度迎接较冷的风,这些身处白垩纪非洲的动物将能更有效地降低体温。
之后有其他研究人员提出反对意见,认为如果棘龙有背帆,它们吸收热量的效率,将高于散发热量的效率。他们认为这些有长神经棘的恐龙,背部具有脂肪构成的隆肉,以储存能量。

许多现代动物的复杂身体结构,在求偶季节时具有吸引异性功能。这些恐龙的帆状物有相当可能性具有吸引求偶功能,类似孔雀的尾巴。斯特莫假设这些恐龙的雄性与雌性拥有不同大小的神经棘。如果属实,这些帆状物可能拥有耀眼的颜色,但这是完全建立于推测上的。

在传统看法里,棘龙被叙述成二足动物,80年代早期之后,它们被认为至少有时可用四足方式行走。这个论点因为发现了重爪龙而得到支持,重爪龙是棘龙的近亲,拥有结实的手臂。
在1997年,Jack Bowman Bailey赞成棘龙为可能四足姿势,这可见于他研究中的新重建图。这个四足步态理论已经过时了,但棘龙科恐龙可能以四足姿态蹲伏。

棘龙步态的骨架模型

棘龙的栖息环境涵盖北非大部分,但只有部分地区被研究过。一个针对摩洛哥恐龙群的研究,认为森诺曼阶的棘龙、鲨齿龙、三角洲奔龙的地理分布横跨整个北非。棘龙生存于当时埃及地区的海岸与潮坪环境,具有红树林,与类似的大型掠食动物共存。

目前仍不确定棘龙主要是陆地掠食动物,还是食鱼性。棘龙拥有延长的嘴部、圆椎状牙齿、以及较高的鼻孔,类似现代鳄鱼,显示它们可能是鱼食性。


棘龙食性的唯一直接证据来自于它们的近亲,居住于欧洲与南美洲的重爪龙。曾在重爪龙的胸腔中,发现了一些鳞齿鱼的鳞片;另一个标本的胃部曾发现幼年禽龙的骨头。另外,在南美洲发现的一个翼龙类化石上,嵌入一个棘龙类的牙齿,显示棘龙类偶尔以这些飞行主龙类为食。
棘龙可能是种无特定目标、多种食物来源的掠食动物,可用白垩纪的大型灰熊来形容它们,平常偏好捕食鱼类,但也寻找许多小型到中型的猎物为食。


在2009年,达鲁·沙索等人对编号MSNM V4047标本进行X射线计算机断层成像,发现棘龙的上颌下颌有疑似感应器官的小孔。达鲁·沙索等人推测,棘龙会将身体浸泡在水中,头部半露出水面,并感应周围的猎物动静,而进行猎食。

在2010年,一项针对棘龙科牙齿的氧同位素研究,显示棘龙是半水生动物。这项研究分析棘龙、重爪龙、激龙、暹罗龙的氧同位素组成,并与同时代的其他兽脚类恐龙(例如鲨齿龙)、乌龟、鳄鱼互 相比较。研究人员发现棘龙牙齿的氧同位素组成,较接近同一地区发现的乌龟、鳄鱼,而不类似同一地区的兽脚类牙齿。研究人员推测棘龙是半水生动物,可在陆 地、水域中生存,以避免与当地的大型兽脚类恐龙、大型水生鳄鱼竞争食物。

棘龙长久以来出现在大众的恐龙书籍里,但直到最近才有足够的棘龙科资讯可以正确地描述棘龙。在过去,棘龙是以直立、背部有帆状物的兽脚亚目形象所重建,直到1955年,艾伯特·拉伯(Albert de Lapparent)与Lavocat建立了一个具影响力的棘龙重建图。

棘龙出现在2001年的电影《侏罗纪公园3》。在电影里,棘龙被描述成比暴龙还要大型、强壮的动物,有着类似异特龙眼前方的角冠,棘龙甚至在一个打斗场景里折断了暴龙的脖子。在现实中,棘龙与暴龙生存在不同大陆,时间也相差了数百万年,因此这两种生物之间不可能发生决斗。
棘龙也出现在电视节目《失落的埃及恐龙》,在节目中棘龙穿越白垩纪埃及的沼泽地。棘龙也出现在探索频道的电视节目《远古巨兽大复活》,节目将棘龙叙述成白垩纪非洲的顶级掠食动物,猎食皱褶龙,并杀死竞争尸体的鲨齿龙、帝鳄。
更多精彩内容,请关注“动物世界”
微信公众号:iltawcom





























 渝公网安备50010702502703号
渝公网安备50010702502703号